Reticulo-endothelial (RES) macrophages are central for the regulation of iron homeostasis. These cells have the ability to handle a high amount of haemoglobin-derived iron, released after senescent red blood cell phagocytosis. Eventually, iron is exported to the bloodstream through the only known iron exporter, ferroportin, and is utilised to support the production of new red blood cells in the bone marrow. This process enables efficient recycling of iron and the maintenance of systemic iron balance.
Beside this role, macrophages are key innate immune cells which exhibit remarkable functional plasticity, reflected in their capacity to integrate diverse signals from the microenvironment and acquire distinct phenotypes. According to a simple dichotomous nomenclature, macrophages are defined as classically activated pro-inflammatory M1 or alternatively activated anti-inflammatory M2.1 M1 macrophages are characterised by a strong bactericidal and bacteriostatic activity and the release of pro-inflammatory cytokines as well as reactive oxygen and nitrogen radicals. Stimuli such as pathogen-associated molecular patterns (e.g. lipopolysaccharide) and inflammatory cytokines (e.g. interferon gamma) drive M1 macrophage polarisation (Figure 1). Typical M1 markers include major histocompatibility complex (MHC) II, CD86, CD14, inducible nitric oxide synthase (iNOS) and inflammatory cytokines, such as interleukin (IL)-1α/β, IL-6, tumour necrosis factor (TNF)α, IL-12, and IL-23. M2 macrophages are mostly involved in tissue remodelling, angiogenesis and immunoregulation. This macrophage subtype is polarised after stimulation with anti-inflammatory cytokines, including IL-4, IL-13 and IL-10, and identified by increased expression of markers such as CD206 (mannose receptor), arginase-1, Ym1, IL-10, transforming growth factor (TGF)-β, CD163 and FIZZ1.
In the last decade, the study of macrophage polarisation has moved into focus, triggered by the in vivo observation of a variety of macrophage subtypes hallmarked by diverse functions and linked to several diseases, including atherosclerosis, cancer, infectious and autoimmune diseases.1–4 Due to their dual role in iron homeostasis and immunity, it has been first hypothesised that macrophage iron handling is affected by cell polarisation.2,5 The acquisition of a specific macrophage phenotype dictates the expression of iron-regulated genes and determines cell iron accumulation and trafficking. M1 macrophages show an iron retention phenotype, hallmarked by low expression of CD163, HO-1 and ferroportin, and high expression of ferritin. In contrast, M2 macrophages show higher iron turnover, associated with increased iron uptake, recycling and export.2 This suggests that cytokines responsible for macrophage polarisation determine the expression of iron-related genes and therefore cell iron handling.2,5
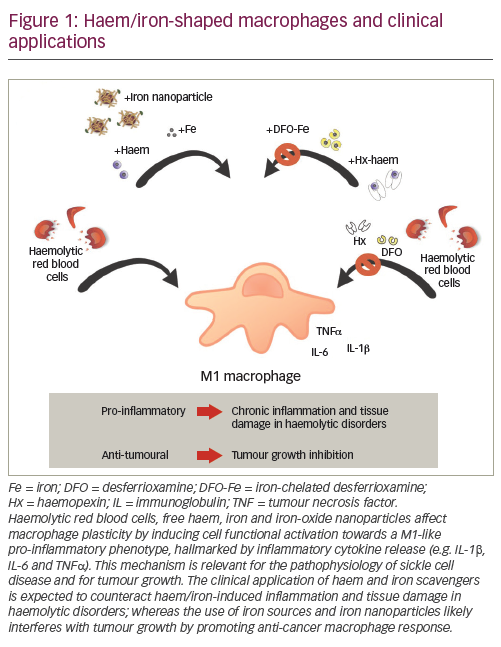
In the second instance, we hypothesised that macrophage iron exposure could directly affect cell polarisation. Interestingly, both in vitro and in vivo, free haem and iron polarise macrophages towards an M1-like pro-inflammatory phenotype.3,6 Haem- and iron-scavenging, through the haem-binding plasma protein haemopexin, or the chelator deferoxamine, prevents this effect (Figure 1). Our findings indicate that iron availability affects macrophage plasticity and determines cell immune effector functions.3 The polarisation programme of macrophages induced by cytokines can be enhanced (M1→M1+) or even shifted (M2→M1) upon further exposure to haem or iron.3 This suggests that iron exerts a dominant effect on macrophage polarisation, inducing a shift toward a proinflammatory phenotype and modulating the phenotypical differences acquired in response to the environmental stimuli.
Importantly haem/iron-driven M1 polarisation has a clear pathophysiological relevance. Sickle cell disease is hallmarked by haemolytic red blood cells and elevated circulating haem levels, which target RES macrophages. In a mouse model of sickle cell disease, iron-loaded hepatic macrophages show an M1-like inflammatory phenotype characterised by increased expression of iNOS, MHCII, CD86, IL-6 and TNFα.4 M1 macrophages promote hepatic damage and fibrosis, and contribute to the chronic inflammatory state which hallmarks sickle cell disease. Consistently, in mouse models of wound healing, acute joint bleeding and spinal cord injury, iron overload induces a pro-inflammatory M1 phenotype, associated with inflammatory cytokine release.7–9 The tumour microenvironment can also present iron-loaded macrophages, as recently shown in a model of lung cancer.4 Those tumour-associated macrophages are enriched at the invasive front of the tumour in haemorrhagic areas and accumulate haem-iron released from haemolytic red blood cells which extravasate from leaky vessels. Interestingly, the presence of higher numbers of iron-loaded tumour-associated macrophages correlated with reduced tumour size in patients with non-small cell lung cancer.4 In the tumour microenvironment, macrophages show a characteristic pro-tumour M2-like phenotype which can be shifted to an anti-tumour M1-like inflammatory phenotype by exposure to haemolytic red blood cells, haem or iron. Iron-elicited inflammatory tumour-associated macrophages show, in fact, the ability to directly kill tumour cells and therefore reduce tumour size.4
Altogether these observations demonstrate that iron-induced pro-inflammatory phenotypic switching of macrophages underlies different pathologic conditions. This suggests that the modulation of macrophage plasticity through iron handling might have therapeutic potential in those diseases. Indeed, haem scavenging via haemopexin administration in mice with sickle cell disease significantly reduces macrophage pro-inflammatory profile, thus ameliorating sickle cell disease inflammatory state.3 Conversely, reprogramming macrophages from the M2 to M1 inflammatory phenotype in the tumour microenvironment could represent a promising anti-tumour therapeutic strategy. In mice, iron delivery in the tumour microenvironment via iron-oxide nanoparticles elicits pro-inflammatory macrophages with tumour-killing properties which significantly delay tumour growth.4,10
Collectively, these observations suggest that iron, in its different forms, has a key role in shaping macrophage phenotype. Data available so far demonstrate the existence of a close interconnection between the immune and the iron handling functions of the macrophage and prove that these activities dynamically influence each other. These findings have potential implications for several pathologic conditions, including: haemolytic diseases, where red blood cell haemolysis and elevated circulating haem might promote a detrimental chronic inflammatory state; infectious diseases, where free haem and iron, released upon cell damage, might boost inflammation and enhance resistance to infections; and tumours, where free haem released in haemorrhagic areas might induce an anti-tumoral inflammatory response. Importantly, current knowledge supports the concept that macrophage plasticity can be manipulated for therapeutic purposes by applying different iron sources or scavengers/chelators according to the desired beneficial effect. In the near future, we expect to learn more in regard to the effect of different iron sources, including red blood cells (e.g. transfusion) and iron preparations (e.g. iron formulation administered intravenously or orally in anaemic patients) on macrophage polarisation. This will help us to discriminate between more and less inflammatory iron sources, with relevance for disease conditions associated with iron/haem overload (sickle cell disease, thalassaemia, paroxysmal nocturnal haemoglobinuria, haemochromatosis) and therapeutic approaches where iron is administered (transfusions, oral/intravenous iron infusion).